| HOME | HELP | FEEDBACK | SUBSCRIPTIONS | ARCHIVE | SEARCH | TABLE OF CONTENTS |
|---|
|
|
||||||||
a Obesity Research Center, St. Luke's–Roosevelt Hospital, Columbia University College of Physicians & Surgeons, New York City
b Department of Pathology, University of Michigan, Ann Arbor
c Department of Biological Science, University of Idaho, Moscow
d Pennington Center for Biomedical Research, Baton Rouge, Louisiana
e Columbia University College of Physicians & Surgeons, New York City
f Jackson Laboratory, Bar Harbor, Maine
g Institute for Behavioral Genetics, University of Colorado, Boulder
David B. Allison, Obesity Research Center, St. Luke's–Roosevelt Hospital, Columbia University College of Physicians and Surgeons, 1090 Amsterdam Avenue, 14th floor, New York, NY 10025 E-mail: dba8{at}Columbia.edu.
| Abstract |
|---|
|
|
|---|
PANEL 5 was given the following charge: Genetic differences among the human population could produce differences in responses to lowered caloric intake or interventions that mimic it. Is there evidence for genetic factors causing variability of the effects of caloric restriction on life span and/or age-related pathologies in different strains or species? Are there additional strains or species that should be tested to probe for these differences? Are there particular loci that should be examined for effects on these responses in experimental animals? What do human data on genetic regulation of metabolism and obesity suggest about variability in responses to sustained low caloric intake or interventions which mimic it, and what additional studies would be most useful?
The panel contended that currently there is very limited information directly addressing these questions, and therefore they focused more attention on what studies could be done than on what studies had been done. In most cases, the panelists believed that certain conceptual issues merited clarification, and some attention was focused on this issue. Although the conceptual issues were similar, the methodological issues and databases available for human and nonhuman models were found to differ markedly. Therefore, human studies and studies of nonhuman model organisms are generally discussed separately in this report.
Before proceeding further, it is worth noting that this report is not intended to be an exhaustive and completely objective review, but simply the report of a panel with widely diverse interests and areas of expertise that met for one day. Such a panel cannot come to agreement on many controversial points nor present a balanced comprehensive review. We incorporate statements from the panel members in the report, so it is inherently contradictory in style and content at points. The result, we hope, is an exploratory, idea-rich (sometimes data-poor), summary of several hours of discussion.
| Background |
|---|
|
|
|---|
Given this background, what do we currently know about the genetic influences on the response to CR in humans? The answer is almost nothing. One set of widely cited studies is that of Bouchard and colleagues (1), which involved examining the genetic control of the response to overfeeding among monozygotic (MZ) twins. These are important studies and show strong genetic influences on the response to overfeeding. However, these do not directly pertain to the issue of CR nor do Bouchard's complementary studies of the genetic influences on response to caloric deficits induced by increased physical activity energy expenditure (2). The only studies the panel was aware of that directly examined the genetic influences on response to CR are those of Hainer and colleagues (3), which evaluated the responses of 14 pairs of obese female MZ twins in Prague who underwent very low-calorie diets in a metabolic ward. Although this study clearly showed evidence of a genetic effect on short-term responses to CR among obese persons, it provides no direct information regarding the genetic influences on the anti-aging or longevity-promoting effects of CR.
The panel acknowledged that there is clear evidence for genetic influences on relative body weight (4), on caloric intake (5)(6), and on age at death (7)(8)(9). These studies suggest that there are likely to be genetic influences on the longevity-promoting response to CR. However, none of these should be confused with evidence regarding the putative genetic influences on the longevity-promoting response to CR.
Caloric Restriction in Rodents
The only treatment that consistently retards aging in mammals is the nutritional manipulation, CR. It extends life spans (LSs) of short-lived rodents if begun early in life (10)(11)(12)(13) and to a lesser extent if begun later in life (14). Knowledge of how CR retards aging processes might suggest clinically useful treatments. Thus, the critical question may not be "Does CR extend human LSs?" but "Can the mechanisms that retard aging in rodents suggest how human aging may be retarded?".
CR does not return laboratory rodents to their "natural state"..-- CR greatly retards reproduction in mice and rats (15). Thus, in the wild, very few calorically restricted rodents would reproduce even once before they were killed. While there may be some circumstances where the delay in reproduction by CR gives a selective advantage (16), rodents in the wild generally cannot exist naturally in CR because they reproduce far faster than do rodents subjected to CR.
CR increases both reproductive and total life spans..-- It can be argued that there is less evolutionary value to increasing the age at which a long-lived animal is capable of reproducing. If CR-induced life extension is an evolutionary adaptation, the primary target could well be the extension of reproductive LS. It has been demonstrated that CR does extend reproductive LS (15)(16)(17). It is tempting to assume that inhibition of the reproductive axis postpones reproductive senescence and leads to extended LS. It is plausible that CR could act as an adaptive mechanism for wild populations in so far as it would permit rodents to survive during times of food shortage and enable them to reproduce at ages older than if they had access to abundant food supply.
Interestingly, there is a strain of mouse, the POSCH-2, that is reported to produce its first litter at 47 weeks of age, and females continue breeding at 3 years of age (18). These animals might be useful in disentangling the effects of CR and delayed reproduction on longevity. However, it should be noted that starting reproduction late may as likely be due to functional defects as to delayed aging, and this is made likely by the fact that these mice contain Robertsonian chromosomes, which affect pairing in meiosis. Moreover, Biddle and colleagues (18) give only very sparse data. Thus, this model, though interesting, is of unknown utility at this time.
Shutting down reproduction is neither necessary nor sufficient for CR-induced life extension. The age at which female mice (unpublished data) and rats are able to bear young is substantially increased by CR. Rats subject to CR, however, may continue to cycle regularly with the same periodicity (17). Thus, the increase in total LS in females generated by CR is not inexorably linked with the reproductive timetable. Additionally, while heavy exercise may shut down reproduction in females, it has not been shown to substantially increase LS. This suggests that simply affecting reproduction in females may not be sufficient to elicit a change in LS in all species. In males, while castration has been demonstrated to confer a slight extension of LS (19), this effect is speculated to be derived from the reduction in early mortality due to dangerous behavior. Thus, a more plausible connection between reproductive scheduling and increased LS in rodents is that some of the same processes underpin control of both the reproductive axis and LS.
CR extends life spans regardless of obesity..-- The relation between LS and body weight or obesity within rodent species is not entirely clear. Allison and Heo (unpublished data) recently analyzed the BIOSURE data (20), which consist of data from 1,200 Wistar rats placed on one of 12 different dietary regimens varying in type and degree of CR and followed for 30 months or until time of death. We used Cox-regression (21) to examine the relationship between body weight and mortality rate. We found that lesser CR and increased body weight were each independently associated with increased mortality rate. Of course, these results must be interpreted with caution because rats were not followed beyond 30 months and, as described below, the effects of CR on mortality rate are also pronounced at late ages. Additionally, mortality was related to weight, which could reflect overall growth, not necessarily adiposity. This stresses the value that widely available in vivo measures of animal body composition would have. Bertrand and colleagues (22) found no relationship between obesity and LS in Fischer 344 (F344) rats. Genetically obese B6 (C57BL/6J)-ob/ob female mouse longevities are extended by CR, although nearly half their body weight is lipid (23)(24)(25). Effects of CR in extending maximum LSs are identical in B6 +/+ or ob/ob females, despite the fourfold higher lipid mass of the latter (23). Thus, CR can increase LSs even in very obese individuals, which may suggest that the benefits of CR are not mediated primarily through reductions in lipid mass. An alternative interpretation is that, as B6 +/+ and ob/ob mice are congenic, meaning that they differ from each other by only a single gene, the similarity in response to CR may simply reflect their near genetic identity rather than addressing the effects of adiposity on the CR response. Thus, the putative effects of weight and adiposity on LS independent of CR remain open to question.
CR does not reduce life-long metabolic function.-- A classical proposed explanation for how CR retards aging is that it reduces metabolic rate. It has been reported that F344 rats consumed equal amounts of oxygen per g lean body mass per day whether on CR or fed ad libitum (26). Consumption of oxygen is consistent, then, with the results of life-long CR studies (27)(28) that have found that restricted rodents consumed more calories/g during their lives than did ad libitum fed controls.
CR does not act only by retarding development.-- If CR is only effective in extending LS when begun before development ceases, this would suggest that it acts by retarding development. In fact, CR begun at 6 months gave about two thirds the extension of median longevities as restricting male F344 rats from weaning at 6 weeks of age (13). CR begun at 12 months gave about one third the extension of longevity as restricting male B10C3HF mice from weaning at 4 weeks of age (14). However, it is clear that CR extends LS even if begun in adult life at least through early middle age (14). In studies by Lipman and colleagues (29)(30), CR begun at 18 and 26 months had no detectable effect given the sample sizes they had available. However, some discussion at the CRCI Advisory Group meeting suggested that there may be models with benefits even at 18 months of age. Clearly, there seems to be a decreasing beneficial effect of CR when initiated at greater ages and, when looking for presumably small effects, sample size and power issues become very important.
Thus, there may be some benefits from CR due to developmental delay. However, rats and mice are sexually mature before 2 months of age, so the substantial benefits at 6 and 12 months are unlikely to be primarily explained by delaying normal processes of development.
Relevant Research in Lower Organisms
The mechanisms mediating increased LS in mammals from CR are unknown, although theories have been proposed (31). However, in C. elegans a genetic pathway with homology to the mammalian insulin/IGF-1 signal transduction pathway recently has been shown to mediate the induction of diapause-arrest in larva; reduction of function mutations in genes of this pathway also increases LS in adults.
The DAF-2 gene is a tyrosine kinase in the insulin receptor family (32) that may signal age-1/daf-23, a PI 3-kinase (33). This signal may antagonize a HNF-3/forkhead type of transcriptional regulator daf-16 (34)(35) that directly regulates the transcription of genes that increase LS in daf-2 and age-1 mutants (36), when kept at temperatures where they do not become dauer larvae. These mutants resist other stresses (37).
Homologies of the DAF-2 worm gene to the mammalian insulin receptor caused Kimura and colleagues (32) to suggest that CR causes a decline in insulin signaling, similar to effects of mutants in DAF-2. Thus, it is noteworthy that in both rodents and primates, CR causes a decline in insulin relative to glucose concentrations (38).
Given that CR in mammals also reduces circulating insulin and IGF-1 (39), this should also diminish components of signaling in the insulin/IGF-1 signal transduction pathway. The mediation of the effect of CR on LS through an insulin/IGF-1 signal transduction pathway may be an evolutionarily conserved mechanism; thus, CR will cause diminished signaling through this pathway with increased LSs. Possibly other treatments that reduce circulating insulin while maintaining normal glucose levels would also increase LS, even if food intake is unaffected.
| Conceptual Issues, Definitions, and Goals |
|---|
|
|
|---|
Second, the panel wished to emphasize the distinction between the suppression of diseases and the retardation of aging. That is, some interventions, including CR, may reduce the risk of certain diseases. Nevertheless, the panel agreed that simply preventing certain diseases (e.g., colon cancer, diabetes mellitus) was not the primary longevity-promoting effect of CR but, rather, that CR appeared to slow some general or fundamental process of aging that affected the rate of development of multiple diseases and of LS itself. It is this latter and more general effect of CR that the panel considered to be of primary interest.
As one indirect piece of evidence for such an effect, it was noted that the survival distribution of humans and most other organisms is much better characterized by a Gompertz than a Weibull distribution. This is noteworthy because the Weibull distribution is the limiting form of the minimum of multiple independent exponentially distributed variables. Thus, if death rates resulted only from "the first random thing that kills you," as some have quipped, then one might expect a Weibull distribution.
The Weibull distribution does characterize much of the human adult LS, but does not effectively characterize the survival distribution at old ages [of course no parametric distribution, including the Gompertz, seems to perfectly characterize the survival curve of any species across the entire life span (40)]. This implies that when we study mortality at old ages we may be more clearly observing the effects of the process of aging itself, whereas in mid-adulthood we may be observing the effects of disease in addition to or instead of aging per se. This point emphasizes the need for long-term studies and studies that include the later components of the LS in studies of CR in which longevity or mortality itself is the variable of interest. For example, in studies validating proposed surrogate measures, it would be important that these measures are predictive of mortality rate at old ages and not just in mid-adulthood. This is similar to the colloquial statement that CR is an intervention that increases "maximum LS" and not just mean LS. In some studies in rodents, CR has little effect on LSs in the first half of the population to die, but increases the LSs in the second half by 20% or more.
Panel 5 agreed that it was important to have a working definition of both CR and the anti-aging effect of CR and offered the following. Caloric Restriction (CR): CR is the chronic consumption of a lesser amount of calories than a comparable "normal" group fed ad libitum where the difference is typically 30 to 40% of energy intake in the control group and micronutrient intake is adequate. The Anti-Aging Effect of CR refers to (a) the reduction of the hazard rate for mortality at multiple points during the LS particularly during later life; and (b) the production of animals that are phenotypically similar to chronologically younger control animals on multiple (but not necessarily all) phenotypes. When begun at weaning, CR postpones reproductive maturity and reproductive senescence.
Panel 5 identified two very broad and reciprocal goals for genetic studies on anti-aging response to CR: (1) To find the genes that influence the longevity response to CR as a means to elucidate mechanisms of action; and (2) To elucidate mechanisms of action as a means of identifying genes moderating the effect of CR on longevity.
| Development of Surrogate Measures |
|---|
|
|
|---|
There have been numerous expositions of criteria for good surrogate measures (41). The panel delineated several criteria that would be useful in evaluating the suitability of a surrogate measure. The first three were thought to be essential and the second three, optional.
Examples of Surrogate Measures
Numerous surrogate measures of aging have been proposed (29)(42). The panel believes it would be useful to offer several examples of better and worse surrogate measures, as there are no uniformly accepted surrogate measures. We therefore provisionally classify possible measures into three groups based on existing evidence. Promising measures are those for which evidence is incomplete, but not incompatible with the stated criteria. As an example, the rate of change in oxidative damage could be one such promising measure. To be an effective measure, the measure of oxidative damage should accumulate with age, the rate of accumulation should be retarded by CR, and the oxidative damage should accumulate more slowly in long-lived species compared with short-lived species.
Evidence on other putative measures is inconsistent with our criteria, and these measures should therefore be dismissed from further consideration as refuted measures. For instance, wound-healing rate was at one time hypothesized to be a useful surrogate measure of aging rate. Although wound-healing rate does decline with age in some species, it does not do so in all species (43). Furthermore, wound-healing for age-matched animals is slower in CR rodents than in AL-fed controls (25). However, there may not be universal agreement on the utility or lack thereof of wound healing as a surrogate measure (44).
Finally, some surrogate measures are of uncertain promise because their relation to our criteria is ambiguous. They therefore need further investigation and clarification. For instance, insulin sensitivity declines with age and its rate of decline is slowed with CR. However, increased physical activity also increases insulin sensitivity, even though it does not retard aging.
Recommendations Regarding Surrogate Measures
The panel made the following recommendations.
• Further development of useful surrogate measures was the highest priority.
• Such development should initially proceed in nonhuman animals and then be evaluated in humans as appropriate.
• Surrogate measures should be sought that meet the criteria specified above.
| The Use of Model Organisms |
|---|
|
|
|---|
A variety of single allele variants that increase longevity are now available in model organisms to help distinguish causal from correlational changes in cellular or systemic processes associated with CR effect. Changes that occur in genetically induced, as well as calorically induced, retardation of aging deserve special attention as pointing to basic mechanisms involved in aging. This involvement can then be evaluated by the development of direct genetic modulation of the process in question. Model organisms that can facilitate study ranging from single-celled organisms to nonhuman primates exist, and all have potential utility (described below).
Aging-Accelerated Versus Aging-Retarded Models
Both aging-accelerated and aging-retarded models are sometimes of interest in aging research. However, animals with apparently accelerated aging are often merely prone to one or a few early pathologies, which are unlikely to be related to mechanisms of CR or basic mechanisms of aging. Gain of function mutants (those that increase the LS normally found in the species) may be more revealing than loss of function mutants. Strains with decelerated aging were believed to be useful in elucidating the plausible physiological mechanisms of CR's effects by showing which physiological patterns were common to both CR animals and animals with mutations that increase LS.
Other Types of Model Organisms
Other types of model systems that can be useful include transgenic animals [for review, see (45)], tissue-specific transgenics, "knock-ins," "knock-outs," and animals with known mutations. Animals created through random mutagenesis or with unusual phenotypes due to naturally occurring polymorphisms can also be useful, as can wild-derived animals (see below for more details).
Recommendations Regarding Model Organisms
• The panel recommended that currently available so-called models of accelerated aging not be used in dissecting mechanisms of CR.
• The panel believed that animal models of decelerated aging were useful and merited further study.
• The panel believed that the continued use of model organisms in CR research was essential and of high priority.
| Use of Candidate Genes |
|---|
|
|
|---|
Characteristics of Good Candidate Genes
The panel concurred that delineating specific candidate genes in this document was not optimal given the rapidity with which information changes in this area. Rather, the panel chose to specify the following criteria for useful candidate genes. Good candidate genes would be those that are known to:
• Affect a biological pathway through which CR is presumed to have effects on longevity; or
• Affect longevity in some species.
Recommendations Regarding Candidate Genes
The panel recommended that:
• The use of candidate genes should be encouraged in general and in human studies in particular.
• Efforts should be made to develop batteries of candidate genes.
• NIH-subsidized genotyping capabilities for these batteries of candidates should be made available.
| Methodological Approaches |
|---|
|
|
|---|
The statistical solution to studying the genetics of the longevity response to CR is to recognize that this can be modeled from a sample of multiple subjects (animals or humans), no one of which has (or can have) its LS measured twice. Put simply, denoting the LS of animals fed AL as La, we can model variations in their LS through the general linear model as:
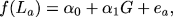 |
 's are parameters to be estimated, G represents the animals' genotype at a locus of interest, and the ea are error terms. Similarly, denoting the LS of animals on CR as Lb, we can write:
's are parameters to be estimated, G represents the animals' genotype at a locus of interest, and the ea are error terms. Similarly, denoting the LS of animals on CR as Lb, we can write:
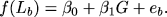 |
Because we are fundamentally interested in the putative effect of G on the difference between La and Lb, we can subtract the first equation from the second, which gives:
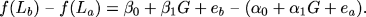 |
Rearranging gives:
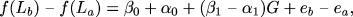 |
1) = 0, that is, whether ß1 = 1. This is just another way of saying that testing for an effect of G on the longevity response to CR is equivalent to testing whether the effect of G on LS is the same for animals fed CR and AL. (Note that this line of reasoning only applies to tests in which one is modeling the first order statistics (means) of LS as a function of genotype or alleles at the locus of interest and not when one is modeling the second order statistics (variances and covariances) as a function of identity by descent (IBD) allele sharing among related individuals at the locus of interest. Therefore, the statistical solution is not identical for typical tests of linkage in humans.) Once it is phrased this way, it is obvious that this can be tested via ordinary interaction tests (46) of the form:
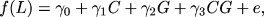 |
 's are parameters to be estimated and C is an indicator function of CR. A statistically significant non-zero value of 3 implies that G affects the longevity-promoting effects of CR. What is important is that, phrased this way, testing the hypothesis of interest can be done with a sample of animals from the same population, some of whom were randomly assigned to CR and some of whom were randomly assigned to AL despite the fact that each animal only lives once. Therefore, the well-developed battery of genetic approaches to testing the effects of putative genes in animals (reviewed below) can be applied to both surrogate measures and LS itself, though the latter may require rather large sample sizes. However, in humans this is not practical and surrogate measures will be essential.
's are parameters to be estimated and C is an indicator function of CR. A statistically significant non-zero value of 3 implies that G affects the longevity-promoting effects of CR. What is important is that, phrased this way, testing the hypothesis of interest can be done with a sample of animals from the same population, some of whom were randomly assigned to CR and some of whom were randomly assigned to AL despite the fact that each animal only lives once. Therefore, the well-developed battery of genetic approaches to testing the effects of putative genes in animals (reviewed below) can be applied to both surrogate measures and LS itself, though the latter may require rather large sample sizes. However, in humans this is not practical and surrogate measures will be essential.
Nonhuman Animals
Inherent value of genetic approaches..--
Genetic approaches have been used repeatedly to understand diverse biological phenomena: oncogenesis, development and the cell cycle, among others, but have only recently been applied to the analysis of organismic aging and senescence. Examples of applications in aging are the genetic dissection of the pathways controlling longevity in the nematode C. elegans and the identification and deciphering of the molecular mechanisms of Alzheimer's disease (AD) through the identification of human genes either causing or predisposing to AD. It is the view of Panel 5 that "Genetics is an important tool for identifying key molecular events that are involved in specifying biological functions." A systematic genetic approach in model systems could lead to an understanding of the phenomenon of CR at the genetic level, including the nature of the molecular interactions underlying increases in maximum LS. The panel recommends initiating a rigorous genetic exploration of increased maximum LSs in both mice and invertebrates.
The panel laid out the following goals for genetic study in nonhuman model organisms.
• Initiate a "rigorous" program to determine how much genetic variation for increased maximum LSs exists in rodents and in invertebrates.
• Initiate a program to define and test candidate genes specifying putative differences in increased maximum LSs in rodents and in invertebrates. These approaches would involve transgenics and directed mutagenesis.
• Initiate studies in large panels of "replicated" segregating populations for mapping genes involved in increased maximum LSs.
• Initiate random mutational studies in rodents and in invertebrates to find genes that specify increased maximum LSs. In particular, it was noted, the assessment of practical surrogate measures should be added to the panel of phenotypes being assessed by the high-throughput mouse mutagenesis programs.
The power of the genetic approach stems from three facts. First, genetic analyses allow the integration of phenomena that must be studied at distinct levels of observation from the molecule to the intact organism. Second, genetics has the power to reveal causality by factors that are not dependent upon the prejudice of the investigator (this does not apply to targeted mutagenesis). Third, genetics allows the unification of observations made in different species based upon sequence similarities of genes in these species. These factors make the genetic approach different from any other experimental manipulation. In the area of CR, the application of a "general genetic" approach could well reveal the causal factors underlying how CR retards aging and allows its genetic dissection. We will briefly elaborate each of these points below.
Point 1. A key problem in understanding the mechanisms of CR is that only one intervention is available, meaning that plus/minus studies are the only ones feasible. Genetic variants that mimic some of the prominent features of CR, especially the increased maximum LSs, offer multiple additional handles for the genetic dissection of the process. Mutants or variants with these effects can be studied at the gene, protein, cellular, and whole-animal level in an effort to understand the molecular details of aging processes. The coupling of these studies at multiple levels to the genetic mutation or variant can lead to insights into the molecular details underlying basic mechanisms of aging.
Point 2. Each investigator is driven by background training and prejudices that suggest which mechanism is likely to be involved in the biological process under analysis. A genetic approach need not be so prejudicial or "hypothesis-driven" in the classical sense of the word. The hypothesis driving the geneticist is that there are genes underlying the CR state, and that variation in these genes either exists or can be generated such that specific outcome variables such as fraction of LS extension, blood glucose levels, levels of advanced glycation endproducts, and corticosterone levels can be examined to see if there is a correlation or even a causal relationship. Selected lines, recombinant inbred (RI) strains, segregating populations or mutants with altered responses to CR reveal which variables are causally involved with life extension and which can be separated without loss of life extension.
Point 3. Finally, evolution has ensured that species evolve by retaining important genetic functions and adapting these functions to novel environmental situations or niches that the species then evolves to occupy. That the increase in maximum LS from CR has been retained over a long period of evolution suggests that it serves some useful function or is so closely involved with some important function that its loss would produce a significant selective disadvantage. Several arguments have been put forth elsewhere as to what such selective advantages could be. The conservation of the response to CR between mammals and invertebrates offers significant potential to use genes discovered in genetically amenable species as starting points for specific explorations in mammals where orthologous genes can be identified and manipulated for possible functionality in mediating CR.
Overview of various genetic strategies..-- Many considerations concerning feasibility and cost effectiveness must be made to ensure the success of such a project. A careful inquiry into the cost effectiveness, the scientific validity, and the likelihood of success of each approach should be carried out. A quick overview of different approaches is given in Table 1 , and potential problems and advantages are shown in Table 2 . These are relevant to model systems and should be contrasted with human genetic approaches such as sib analysis. These are genetic strategies and do not refer to particulars about what surrogate measures should be used or how these surrogates should be established.
|
|
Problems in diallele analysis, selective breeding, and quantitative trait loci (QTL) mapping..-- The most significant distinction between the first three of the approaches listed in Table 1 and the last two is based on the origin of the genetic variation being analyzed. The first three approaches rely on genetic variation already within the population being studied and cannot identify alleles that are genetically fixed. Thus, it is vital to use strains descended from populations that vary in critical life history phenotypes, such as reproductive rates and longevities. Strains recently derived from the wild and even wild-trapped progenitors may provide differences in alleles that have become fixed with domestication. Alleles with significant negative effects on fitness components of life history traits will be found only at very low frequencies, so that the genetic variation necessary for effective genetic analysis using these approaches will be lacking.
This is not to say that alleles with effects on CR will not be polymorphic; rather, that those genes that are polymorphic will not be the major genes specifying CR characteristics in the species if there is no variation in the species. Moreover, if CR is really genetically conserved across species, it must be because it confers selective advantages in almost all species. The frequency of defective alleles segregating in the population would be kept low and thus be difficult to detect using only preexisting genetic variation. The mode of action of genetically fixed alleles or genes may be most informative in understanding the genetic specification of CR, if these alleles determine the rates and mechanisms of aging in each species. Such genes may be identified by mutational studies, as has been demonstrated both by the work in the nematode and Drosophila.
Determining which rodent strains to test..-- —Naively crossing short- and long-lived strains may identify trivial factors, as there may be an indefinite number of trivial explanations as to why LSs are reduced in a particular strain. Most probably it will be a particular pathology. An alternative approach would be to cross highly divergent strains (different subspecies and interfertile species) in the hope that there are significant genetic differences in their key life history alleles.
Another approach is to cross animals evolved under different kinds of selective pressures. One of the most important aspects of their habitat is how likely they are to encounter the problem of fluctuating resource availability:
• Seasonal versus tropical
• Warm versus cold
• High versus low predation levels
• Selection for large versus small size
To look for QTLs involved in CR one must find early phenotypes (metabolic, endocrine, etc.) or surrogate measures. Ideally these should map to the same chromosomal location as QTLs that extend maximum LSs. It may be impractical to use life extension as the only phenotype because each experiment will require years to conduct. In addition, to achieve a reasonable map resolution (to proceed with candidate genes) with a quantitative trait, many hundreds of animals may be needed; this is extremely expensive for a life-long experiment.
Diallele Analyses..-- In this approach, a panel of inbred strains are crossed among themselves and the resulting series of F1 hybrids are studied for the CR phenotype. The major advantage of this approach is that it is fast and very replicable. The fact that large numbers of genetically identical animals can be studied for various traits is important, as well. The major disadvantages are two. First, there is very little ability to find individual genes underlying the genetic variation, so the results serve best to define the degree of genetic variation in the progenitor strains. Panel 5 recommends performing these crosses in mice to confirm and estimate the extent of genetic variation in CR.
Selective breeding..-- The major advantages of selective breeding are in establishing that a simple additive genetic component for CR exists and in providing lines (non-inbred) of mice that vary dramatically in their response to CR. The major disadvantages are the long period of time involved in their construction and the inability of selective breeding alone to uncover individual genes that are responsible for the differences in the CR phenotype. The latter problem can be overcome by the generation of segregating populations by crossing lines with different phenotypes. The mapping of genes in such crosses is possible, but difficult.
QTL mapping.-- This approach has most of the advantages of both diallele analysis and selection as well as those of random mutagenesis. It shares the problem of utilizing preexisting variation, which is the major disadvantage of this approach. It provides estimates of size of genetic variation and it allows the localizing of these variants to chromosomal regions. A spate of recent publications have demonstrated that genes specifying each QTL can be identified, and this will only get easier as the mouse genome project reaches maturity. The major disadvantage with some QTL mapping approaches (e.g., F2 mapping) is the lack of replication at the segregating individual level because each is a genetically distinct individual.
This problem may be specially acute in the CR phenotype because a single individual cannot experience CR and be its control. This problem can be overcome by using a "stable segregating population," for example by studying RI strains made by inbreeding F2 sibs from crosses between two parental inbred strains. Such inbreeding fixes the genotypes within each RI strain whose genotype is a unique random set of alleles, half from each of the two progenitor strains. An indefinitely large population of genetically identical individuals is available from any RI strain; thus, any number of distinct and even interfering genotypes can be studied. RIs offer the potential for destructive analyses of the CR phenotype. A major disadvantage of RIs is limited power, as it is extremely expensive to maintain large sets of RI strains. If funds are available, this can be overcome by studying large panels of RIs, which can detect QTLs explaining only a small percentage of the genetic variance.
Probably the major disadvantage of RI lines is that only genes with different alleles in the two progenitor strains can be defined. Unfortunately, RI lines cannot easily be prepared between inbred progenitor strains that are too diverse. Thus, RI lines cannot be used to analyze the full range of variation in life history regulating genes available in the mouse.
Mutagenesis..-- A systematic study of longevity (and age-sensitive traits) in mutant strains of mice (including artificial mutants made by transgenesis or gene deletion methods) that mimic some of the prominent features of the CR state is of great importance. For example, it is striking that the only single-gene mutants known to prolong LS in mammals, the dw and df dwarfing mutants, produce animals that share a multitude of features with CR mice, including small stature, low growth hormone levels, low thyroid hormone levels, and possibly alterations in glucose/insulin balance (47)(48)(49). It is equally informative that these mutants differ in provocative ways from the CR animals; the dw and df mice are, for example, remarkably obese in late life, unlike CR mice.
Construction of animals that have partial deficits, or deficits limited to specific cell types, or whose deficit can be regulated by attachment to a signal-sensitive promoter complex, should in the long run prove more useful than mutants with completely null phenotypes. In the current context the emphasis would be on determining the ways in which these mutants reproduce (or fail to reproduce) key features of the CR paradigm, toward the goal of defining the elements of the CR state most important for its effects on late-life disease and on longevity.
Advantages and Disadvantages of Random Mutagenesis..-- The major advantage is that the entire genome becomes available for the identification of genes specifying CR. The study is not limited to those genes that are polymorphic at reasonable frequencies, for any gene that is consistent with viability can be detected and studied. These approaches are costly, but, because several random mutagenesis studies in mice are underway, it would be important to include assessments of surrogate measures in these studies, so as to ascertain whether or not there are significant alterations in the status of these surrogate measures. If such changes are found, then the "mutation" would need to be stabilized and confirmed and then it could be used to extend the CR phenotype and to identify novel genes that are involved in the specification of CR. Because this requires coordination with studies now underway, it would be useful were it done promptly.
Advantages and disadvantages of targeted mutagenesis..-- Targeted knockouts and transgenics overexpressing a gene of interest are genetic models that have been available for the last decade or longer and that can be applied to studies of the genetics of CR. Such models offer the potential to test specific hypotheses using the full range of genetic interventions and thus can be very relevant to the study of CR. In addition, the use of regulated and inducible constructs as well as tissue-specific expression and "knock in" technologies allow a full range of genetic variation to be studied to see if there is relevance to the CR phenotype. While this is an extremely powerful approach to testing the importance of particular genes, it has the great disadvantage that it cannot detect new genes that might lead to new mechanisms and ideas.
Humans
Given surrogate measures to work with, a well-elaborated battery of techniques for finding chromosomal regions containing specific genes and alleles with effects on quantitative phenotypes in humans exists and continues to be developed (50)(51). Similarly, techniques for studying gene function and expression exist and continue to be developed (52). The reader is referred to referenced sources for details on these methods.
One general class of techniques is meiotic mapping, which includes such techniques as linkage analysis, association analyses, and joint tests of linkage and association (50). These approaches may be useful in general and may be especially useful in certain unique populations. A population that may be of interest in this regard is Sardinians, who (a) have a relatively high degree of genetic isolation, homogeneity, and disequilibrium (53), and (b) have an unusually high prevalence of exceptionally thin people (54).
Expression studies were also judged to be of potential value. For example, cDNA microarrays can be used to quantify gene expression for tens of thousands of genes simultaneously by fluorescently labeling cRNA or cDNA from tissues that differ in ways likely to be associated with revealing differences in gene expression. For detection of genes related to the anti-aging effects of CR, such arrays might be used to look at expression differences in tissues from humans before and after CR. Illustrative is a recent experiment with fibroblasts exposed to serum where genes found to have coordinate expression were related by having comparable functional properties (55). Thus, genes related to response pathways can be identified without prior knowledge of their existence or relevance. Genes or candidate ESTs (expressed sequence tags) identified in this manner could be vetted by confirmation of transpecies conservation and sequence comparisons among these genes. These genes would also be examined in CR and exercised animal models looking for CR-specific differential expression in organs possibly beneficially affected by CR but not exercise.
Finally, in the human realm, the panel agreed that there was great value in twin studies. By exposing twin pairs to CR (3), one can determine which CR-induced changes in putative surrogate measures are most heritable and, thereby, provide an additional validation of a surrogate measure that is presumed to be involved in the pathways from genotype to CR-longevity response.
Panel 5 concurred that human studies were likely to be difficult but important and wished to emphasize several key points. For human studies to be practical it was believed that:
• The availability of surrogate measures was essential because of the impracticality of assigning humans to CR and observing their LS.
• The availability of candidate genes was very important. This is because tests of linkage and, especially, association are likely to be substantially more powerful when a candidate gene is available than when not, and low power continues to be among the greatest obstacles that human gene mappers face.
• Phenotypes should be measured with the utmost rigor in controlled settings.
• The integrity of the independent variable should be rigorously achieved through careful experimental control. The panel felt strongly that uncontrolled observational studies that did not directly manipulate CR were of more limited value.
| Overall Recommendations |
|---|
|
|
|---|
1. Do not use currently available models of accelerated aging to study the genetics of the longevity response to CR. The panel believed that, were true models of accelerated aging available, such models would be useful. However, they did not believe that currently available models of accelerated aging merited the appellation and therefore did not believe their use in studying the genetics of the longevity response to CR was merited at this time.
2. Do not treat the concepts of thinness, leanness, negative energy balance, and variations in physical activity as though they were synonymous with or indicators of CR. The panel concurred strongly that the concepts of thinness, leanness, negative energy balance, and variations in physical activity are distinct from the concept of CR and that such concepts should not be conflated in the design and interpretation of CR related-studies.
3. Establish centers for normative LS studies of multiple strains and animals with defined genotypes at particular loci. The panel endorsed the establishment of centers for normative LS studies of multiple strains and animals with defined genotypes at particular loci. It would be useful if such centers also had the capacity to measure key putative surrogate measures.
4. Subsidize centers for high-throughput genotyping and use of advanced molecular genetic techniques. Several panel members indicated that it would be very useful if NIH-subsidized centers were available for high-throughput genotyping and use of advanced molecular genetic techniques (e.g., chip-based technologies) for CR investigators.
5. Establish and support a computer-searchable database with lifetable and aging data. A computer-searchable database should be supported with lifetable and aging data for multiple strains under multiple defined conditions. The exact data to be included in such a database merit further discussion.
6. Use animal studies to develop surrogate measures. With respect to animal studies, they were believed to be essential to finding genes involved in the anti-aging effects of CR, and the use and development of surrogate measures in such animals is of high priority.
7. Emphasize human studies with experimental manipulations and/or rigorous phenotyping over observational studies with weak phenotypic measurements. Human studies involving rigorous measurement of surrogate measures and rigorous experimental control of the independent variable were thought to be of high value. Human studies involving extensive reliance on self-reported data were believed to be of lesser value in this topic of study.
| References |
|---|
|
|
|---|
This article has been cited by other articles: (Search Google Scholar for Other Citing Articles)
 |
J. E. Morley Editorial: Hot Topics in Geriatrics J. Gerontol. A Biol. Sci. Med. Sci., January 1, 2003; 58(1): M30 - 36. [Full Text] [PDF] |
||||
|
|
J. E. Morley Editorial: Citations, Impact Factor, and the Journal J. Gerontol. A Biol. Sci. Med. Sci., December 1, 2002; 57(12): M765 - 769. [Full Text] [PDF] |
||||
|
|
A. Fisher and J. E. Morley Editorial: Antiaging Medicine: The Good, the Bad, and the Ugly J. Gerontol. A Biol. Sci. Med. Sci., October 1, 2002; 57(10): M636 - 639. [Full Text] [PDF] |
||||
|
|
J. E. Morley Editorial: Drugs, Aging, and the Future J. Gerontol. A Biol. Sci. Med. Sci., January 1, 2002; 57(1): M2 - 6. [Full Text] [PDF] |
||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| HOME | HELP | FEEDBACK | SUBSCRIPTIONS | ARCHIVE | SEARCH | TABLE OF CONTENTS |
|---|
| All GSA journals | The Gerontologist |
| Journals of Gerontology Series B: Psychological Sciences and Social Sciences | |